Cell adhesion & the integrity of plant organs
Plant cells have an extended cell surface beyond the plasma membrane - the cell wall - and this has a great impact on the modes of cell development. At cytokinesis one cell becomes separated into two as a partition, known as a cell plate, is constructed across the middle of the cell. New primary cell wall is then assembled at either side of this cell plate and the cell plate becomes an intercellular matrix or middle lamella (see scheme below). Such cells generally have each other as fixed neighbours for life there is generally no slipping, sliding or gliding of adjacent cells during growth. However, cells can separate and plant cell separation can be broadly placed into three types.
- Separation of two large masses of plant cells subsequent to the differentiation of specific separating cell layers. This is the process that results in the loss of organs such as leaves and fruit by abscission.
- The detachment of single cells from a larger mass of coherent tissue as seen at root caps and inner seed coats (Willats et al., 2004).
- A limited separation of adjacent cells in tissues such as parenchyma resulting in intercellular spaces as seen most typically in parenchyma tissues when viewed in transverse section (see below).
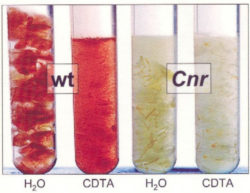
Contrasting cell separation characteristics of ripe tomato pericarp from wild type and the cell adhesion mutant Cnr when placed in water or the calcium ion chelator CDTA (Orfila et al., 2001).
The intercellular matrix is thought to consist largely of pectic polymers and to be free of cellulose - but a detailed description of its components has been hard to achieve. What factors control separation and how are the specific regions of cell walls disassembled? Understanding of these processes is fragmentary although it is well established that polygalacturonase enzymes act during abscission processes. The cell wall polymers that are modified during the loss of cell adhesion are pectic polymers. The homogalacturonan (HG) component has a highly variable patterning of methyl-esterification and it can be cross-linked by calcium ions through its unesterified domains. Blocks of unesterified HG, recognized by phage display monoclonal antibody PAM1, often occur specifically in cell walls that surround intercellular spaces (Willats et al. 1999; Manfield et al., 2005). We have identified a partially methyl-esterified HG epitope, bound by antibody LM7, that occurs specifically at the separating corners of developing intercellular spaces in all parenchyma systems (Willats et al., 2001).
In certain tissues, such as the ripe tomato fruit pericarp, calcium ion cross-linked de-esterified HG is a major adhesive bond and placing tissue in a calcium chelator can rapidly lead to a complete separation of cells. However, this is rarely the case in actively growing organs. The pectic polymers rhamnogalacturonan-I (e.g. Orfila et al., 2001) and rhamnogalacturonan-II have also been implicated in cell adhesion processes. The latter forms links with boron through its apiose residues and this appears to be a major factor in individual cell wall integrity but also possibly contributing to links between adjacent cell walls.
Plant cells arise in meristems attached to neighbours and once separated do not readily attach to each other or aggregate. However, separate plant cells can actively adhere as seen at graft junctions and during carpel development. It appears that the lack of aggregation may be a defensive response, a key aspect of reproduction strategies and also of the interaction with other species and plant parasites. That cell aggregation has to be actively prevented is suggested by analyses of mutations that allow cells to freely aggregate and these analyses indicate that lipid molecules may have a role in this (Jarvis et al., 2003).
A role for hemicellulose polysaccharides?
Recent work with tomato fruit pericarp parenchyma has resulted in methods to isolate cells enabling direct analysis of polysaccharide epitopes at adhesive surfaces. This study indicates that the LM15 xyloglucan epitope has patterns of occurrence in relation to cell adhesion in this system. Moreover, unlike the LM7 HG epitope, these patterns are lost or reduced both in Cnr and during fruit ripening. (Ordaz-Ortiz et al. 2009).
Scheme showing formation of intercellular spaces in parenchyma systems
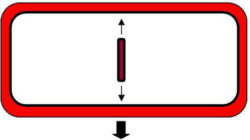 At cytokinesis the cell plate grows from the centre of the cell
At cytokinesis the cell plate grows from the centre of the cell
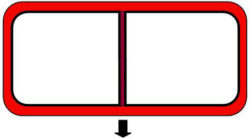 The membrane-bound cell plate partitions the parent cell producing two daughter cells
The membrane-bound cell plate partitions the parent cell producing two daughter cells
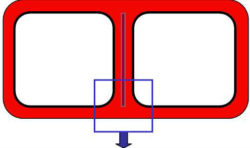 As a cell grows primary wall is deposited on both sides of the cell plate, which becomes the middle lamella
As a cell grows primary wall is deposited on both sides of the cell plate, which becomes the middle lamella
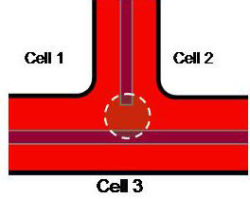 A region of the primary cell wall (enclosed by dotted circle) between the new middle lamella and the original middle lamella of the parent cell is dismantled to allow...
A region of the primary cell wall (enclosed by dotted circle) between the new middle lamella and the original middle lamella of the parent cell is dismantled to allow...
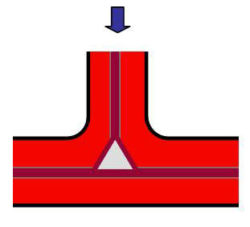 ...the old and new middle lamellae to link up to form an intercellular space between three cells.
...the old and new middle lamellae to link up to form an intercellular space between three cells.